
|
|
|
{Page 21}
Department of Biology, University of Central Oklahoma, Edmond, OK 73034
Oklahoma Water Resources Board, Oklahoma City, OK 73118
Boehler Lake is a 2.5 ha, dystrophic beaver pond. Samples of benthic and eponic fauna were collected over four seasons from April 1994 to January 1995. Four sampling stations representive of different habitats in the pond produced 5,117 individuals and 99 taxa represented by 1 family of nematodes, 3 families of annelids, 3 families of mollusks, 4 families of crustaceans, and 29 families of insects. Dipterans composed 86% of all individuals of insects found. Detritivors and predators dominated the collections. Chaoborus punctipennis and several taxa of Chironomidae were present at all stations throughout the study period. Physicochemical sampling of the pond revealed generally low dissolved oxygen concentrations, and predominantly acidic conditions during the study period. Based on a light/dark bottle study, gross primary productivity was calculated to be -0.10 mg/L oxygen/day. The extensive peat mat, shallow basin, and apparent collapse of the trophic structure indicate that Boehler Lake is in its final stages of succession as an aquatic habitat. ©2001 Oklahoma Academy of Science
| Introduction | Methods | Results and Discussion | References | Top of Page | Table of Contents | Home |
The Boehler Seeps area of southeast Atoka County is a unique natural area of Oklahoma. It results from several geological and biological factors that have occurred nowhere else in the state. Boehler Seeps and Sandhills Nature Preserve was formed on a bed of fine- to medium-grained sands known as the Antlers Formation. These sands, resting upon much older strata that slopes toward the surface, created a water table that remains relatively constant at 5 m (1). These combined elements provide an upwelling effect that feeds water to the surface, and a small lake has formed behind a beaver dam. At the time of this study, Boehler Lake had a surface area of approximately 2.5 ha and a maximum depth of 1.6 m.
Most studies of benthic macro-invertebrate communities in lentic environments of Oklahoma have been limited to reservoirs (2-9). These artificial environments are probably quite different from a small lake formed as a result of springs or seeps being contained by a beaver dam. The purposes of this project were to establish baseline data regarding composition of the benthic and eponic fauna, determine annual ranges of selected physicochemical parameters, and describe ecological trends occurring in Boehler Lake.
| Introduction | Methods | Results and Discussion | References | Top of Page | Table of Contents | Home |
Four sampling stations representative of the major habitats were established in Boehler Lake. Each station was chosen on the basis of its benthic substrate, vegetation type (emergent, submerged, or floating), variety of vegetation present, and the general location within the lake. Each site was visited, and samples of macroinvertebrates were collected, seasonally (April 1994, July 1994, October 1994, and January 1995) during the study period. In addition, during each collection period, water samples were taken to analyze for selected physicochemical conditions.
Field testing of water samples included measurements of dissolved oxygen (DO) concentration (modified Winkler method), temperature (centigrade thermometer), pH
{Page 22}
(Hanna pHep pocket pH meter, ), alkalinity [sulfuric acid method as per Hach Chemical Company protocols (10)], and Secchi depth (20 cm dia. Secchi disc). In the laboratory, testing for nitrate concentration (cadmium reduction method), nitrate concentration (diazotization method), ammonia concentration (Nessler's method), and turbidity was conducted. These laboratory analyses were done with the use Hach Chemical Company reagents and 1992 protocols specified for use with a Bausch and Lomb spectrophotometer model 20. Specific conductance (Oakton conductivity meter) was also measured in the laboratory. A 24-h productivity investigation was conducted in the field by using light/dark bottles (11) during October 1994.
Four benthic samples were taken at each site during each collecting period by using a petite Ponar grab. Each sample was washed in a 250 µm mesh sieve bucket, placed into separate jars, and preserved in the field with a mixture of formalin and rose bengal dye. These samples were transported to the laboratory for washing, sorting, identification, and enumeration.
Eponic sampling involved cutting portions of the vegetation to be studied. These samples were carefully removed from the water to avoid dislodging any macro-invertebrates before being placed in a sieve bucket and treated in the same manner as the benthic samples. After the macro-invertebrates had been removed from the sample, the vegetation was dried, weighed, and its surface area estimated so different samples could be compared (12).
Shannon's index of diversity (13) was used to calculate species diversity of the petite Ponar samples. Sorenson's index of similarity (14) was used to compare faunal similarity and infer community overlap, based on both benthic and eponic samples.
| Introduction | Methods | Results and Discussion | References | Top of Page | Table of Contents | Home |
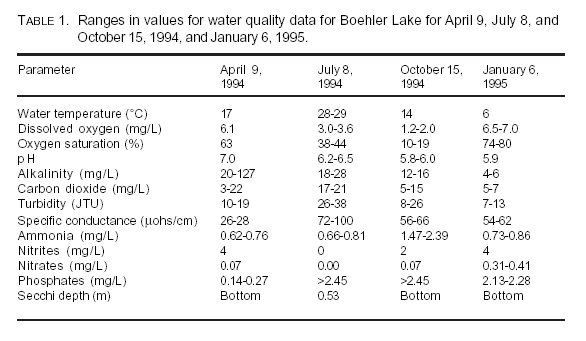
Physicochemical Conditions: The temperature of Boehler Lake reflected the seasonal changes of the area. Temperatures ranged from 6°C in January to 29°C in July (Table 1).
Dissolved oxygen concentrations ranged from 1.2 mg/L during October to 7.0 mg/L during January (Table 1). Although DO concentrations appear to be inversely related to water temperature, it is likely that the level of decomposer activity versus photosynthetic activity was also a contributing factor in the seasonal changes observed in DO concentrations. The length of daylight during each season would affect the rate of photosynthesis while temperature would affect the rate of decomposition of the peat
{Page 23}
mat. It should also be noted that no stratification of DO was observed, probably due to the shallow depth of Boehler Lake.
Productivity of Boehler Lake was measured in October by submerging light/dark bottles for 24 h (11). The gross primary productivity was determined to be -0.10 mg/L oxygen/day during that period. This negative value implied that Boehler Lake was not maintaining enough oxygen near the bottom to support aerobic decomposition at that time. Because this method assumes conditions in the bottles are representative of the rest of the lake, it is hypothesized oxygen is diffusing from the atmosphere to sustain decomposition activity.
These data suggest oxygen is a limiting factor in the lake during the warmer period of the year. During the cooler months, it appears oxygen levels are suitable to support many benthic invertebrates. Cooler water temperatures would also decrease the oxygen demand from decomposition. Theinemann (15) established that oxygen demand exhibited by decomposition is a crucial factor affecting DO concentrations, especially in benthic regions. Furthermore, cooler waters have the capacity to absorb more atmospheric oxygen than do warmer waters (11).
Values for pH ranged from 5.8 during October to 7.0 during April (Table 1). It is suspected this is indicative of an annual cyclic tendency between acidic and neutral conditions within the lake. The acidic nature of this lake was probably due to the addition of allochthanous material, primarily leaf debris and Sphagnum debris, along with autochthanous debris naturally increasing during the fall and winter from primary producer die-off. This combined with the increased decomposer activity occurring in the spring may have acted to reduce the pH further. As decomposition exceeds photosynthetic activity, excess carbonic acid, which reinforces the acidification, is formed. The reduction in pH is probably associated with the accumulation of large amounts of decomposing plant debris. Cole (11) described how coarse organic particulates begin to accumulate because decomposition of cellulose may be reduced as pH approaches 5.0.
Alkalinity concentrations ranged from 4 mg/L in January to 127 mg/L in April (Table 1). These data support that Boehler Lake is acidic and has low buffering capacity. This may also leave the water susceptible to continued acidification as additional respiration products are released during decomposition. Carbonate impoverishment has been reported in Carolina bays (16) and in bogs of the Canadian shield (11). Concentrations of free carbon dioxide were generally low, ranging from 3 mg/L to 22 mg/L (Table 1).
Turbidity measurements varied from 7 JTUs in January to 38 JTUs in July (Table 1). These changes are most likely due to seasonal changes in the sizes of phytoplankton populations or the amount of suspended solids in the water column. However, no further tests were undertaken to determine the causes of turbidity.
Values of specific conductance were generally low, ranging from 26 µmohs/cm during April to 100 mmohs/cm during July.
Ammonia concentrations ranged from 0.62 mg/L in April to 2.39 mg/L in October (Table 1). Cole (11) reported the major source of ammonia in lakes was the bacterial and autolytic activity in detritus. Given the shallow nature of Boehler Lake and the low oxygen conditions caused by decomposition of the peat mat, it is possible ammonia could easily diffuse throughout much of the lake.
Nitrite values ranged from 0 mg/L in July and October to 4 mg/L in April and January, while nitrate values ranged from 0.00 mg/L in July to 0.41 mg/L in January. Because nitrification is an aerobic process, the reduced DO concentrations may be partly responsible for the lack of ammonia oxidation to nitrite and subsequently nitrate. In addition, the extensive amount of vegetation in Boehler Lake may be absorbing nitrates as they are synthesized or released during the growing season and releasing them as die-back occurs during winter.
Phosphate concentrations ranged from 0.14 mg/L in April to greater than 2.45 mg/L in both July and October (Table 1). These higher concentrations occurred during the warm months when plant phosphate utilization should be greatest and were unexpected. DeVito and Dillion (17) found that
{Page 24}
input of forest leaf litter can account for phosphorus accumulation in benthic waters. In addition, they found the activities of beavers contribute to total phosphorus concentrations in lakes where they are present. These explanations may account for the higher phosphate concentrations.
The presence of such high phosphate levels may have contributed to the decreased Secchi depths observed during July (Table 1). The high phosphate concentration probably contributed to higher epilimnetic algal growth, which decreased light penetration and subsequently DO concentration near the lake bottom. This depressed DO concentration would then slow decomposition, increase retention of detritus, and slow nutrient cycling.
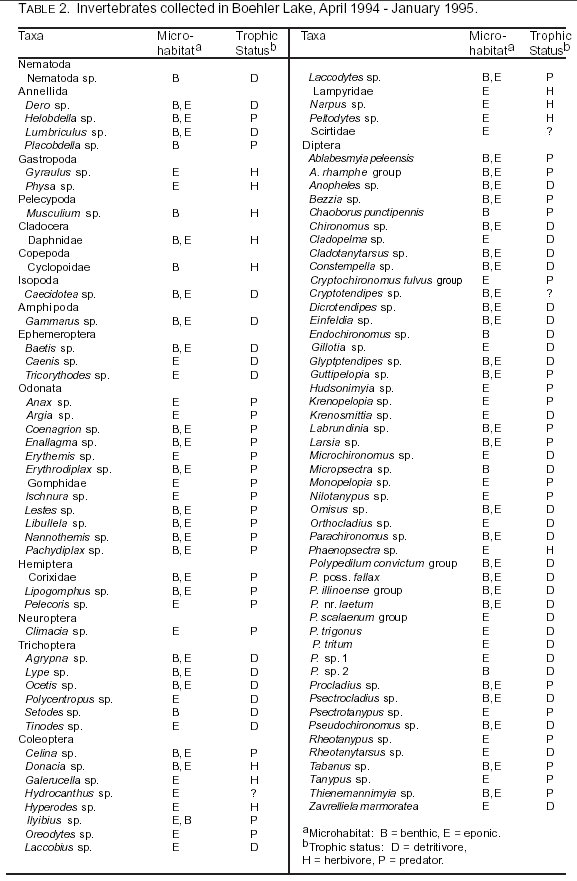
Benthic and Eponic Fauna: During the course of this investigation, 5,117 individual macroinvertebrates representing 99 taxa were collected and identified. Of these, 286 were crustaceans, and 3,692 were insects, representing 4 families and 29 families respectively. The remaining individuals were nematodes, annelids, and mollusks (Table 2).
Several groups of crustaceans were present in Boehler Lake. Cladocerans constituted only 0.5% of the fauna. Amphipods, which composed 4.3% of all fauna, were most abundant in the vegetation and present year round at most stations. Isopods contributed only 0.5% of all fauna. Because the cyclopoid copepods collected during July are usually planktonic, it is assumed they were accidentally captured as the grab was lowered to the bottom.
Ephemeropterans constituted 1.2% of the insect fauna. Odonates formed a higher proportion of the insect community, constituting 7.6% of the insects. Hemipterans made up only 1.7%, trichopterans made up only 1.3%, and coleopterans made up 2.9% of the insect community. None of these groups were frequently encountered nor abundant in Boehler Lake. However, dipterans were extremely abundant, making up 86% of the insects collected.
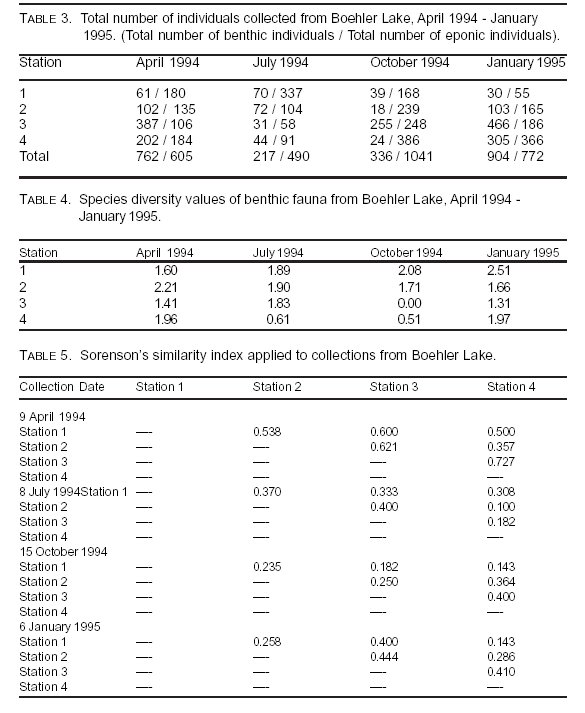
There were marked seasonal differences in the total number of individuals of all fauna at each station and in benthic and eponic samples (Table 3). Changes in numbers of individuals present are most evident
between benthic and eponic fauna from October to January. These changes in community sizes were similar to those documented by Craven and Brown (2) in a deeper, oligotrophic reservoir. In both cases, population sizes generally increased as seasonal water temperatures cooled and depth increased. No migration from the sediments toward the aquatic vegetation was indicated in the data. It is hypothesized the changes in numbers of individuals seen at all stations are most likely associated with the emergence of adult insects. All stations, except Station 1, showed decreases in numbers of individuals during July, which may have corresponded to an emergence period. The number of individuals at Station 1 remained high in July and did not drop until the October collection. All benthic populations decreased to a much greater extent than did the eponic fauna during July. Later, between July and October, the benthic populations increased by 58%, most likely because of the hatching of eggs during that time. Eponic populations also increased slightly during that late summer period.
Separate diversity indexes from each station and each substrate type were calculated (Table 4). Benthic species diversity decreased at Station 4 during the warmer months. Station 2 diversities decreased slightly, while Station 1 diversities increased throughout the duration of the project. Diversity values at Station 3 fluctuated during the study period.
Although all stations maintained diversities below 3.0 for the entire year, reflective of the dystrophic nature of Boehler Lake, Station 3 benthic diversity dropped to zero in October when Chaoborus punctipennis was the only taxon found. Station 4 also displayed a drop in diversity, but its decrease came during the July and October collections. Values for July and October were 0.61 and 0.51, respectively. C. punctipennis also dominated in those samples.
The decrease in diversity values occurring at Station 3 during October and Station 4 for July and October may have resulted from the emergence of several groups of aquatic insects at those stations prior to sampling. In contrast, the decrease at Stations 3 and 4 and the lack of a similar decrease at
{Page 25}
{Page 26}
Stations 1 and 2 may indicate some movement into shallower parts of the lake during the hottest part of the year. If this is the case, then emergence events may occur enmass throughout the summer. Movement into shallower water may have masked emergence events at Station 1. The increases in diversity at Stations 1, 3, and 4 in the January collections may indicate the repopulation of taxa through immigration and the hatching of eggs laid by the previous generation following emergence.
Over the period of this investigation, C. punctipennis and chironomids dominated separate vertical strata. C. punctipennis was found exclusively in the benthos, which is consistent with the habitat preference cited by Merritt and Cummins (18). Because the samples were only collected in the daylight, any diurnal migration into the water column by the chaoborids would have been missed. Chironomids were predominantly found in the vegetation (75% of all individuals) throughout the year. This is in contrast to the benthic habitat preference of most chironomids stated by Merritt and Cummins (18). There was an increase in benthic chironomid populations during January and April, possibly indicating an overwintering strategy for some larvae.
Although a large amount of aquatic vegetation was present, examination of taxa by trophic status reveals very few herbivores existed in the lake. Detritivores and predators dominated at all stations and during all collecting periods (Table 2).
Populations of most taxa were generally too small in number to use as indicators of emergence. Any change in the numbers of a small population would not definitively indicate the occurrence of an emergence event. However, because of their constant presence and marked changes in population sizes, six dipteran taxa were examined for possible emergence patterns. These taxa included Ablabesmyia sp., Bezzia sp., C. punctipennis, Chironomus sp., Larsia sp., and Polypedilum sp. Ablabesmyia sp. appeared to emerge between April and July, as did Bezzia sp., C. punctipennis, Chironomus sp., and Larsia sp. Their population sizes in the July collections were 94, 93, 75, 83, and 62% lower respectively than the April collections.
Ablabesmyia sp. and C. punctipennis showed rebounds in population sizes in the October collections Populations of Chironomus sp. and Bezzia sp. remained low through at least October before showing an indication of a rebound. Repopulation, presumably from the eggs of resident and emigrant adults became apparent as all populations except Polypedilum sp. and Larsia sp. began to return toward previous levels between October and January. Polypedilum sp. appeared to emerge in the late fall or early winter because population sizes were high in the October collections, but greatly reduced in the January collections. Examination of the benthic diversities and the population sizes of the selected taxa seem to offer corroborating evidence of emergence. Furthermore, examination of the distributions of eponic organisms did not seem to support any change in the vertical distribution of these taxa.
Some changes in the population sizes of eponic taxa occurred during this investigation, but this did not form any sort of distinct trend. Examination of the numbers of taxa found in benthic samples as opposed to specific vegetation (eponic) types also did not demonstrate conclusive trends. Many taxa were found only in vegetation samples. Some taxa that were captured in benthic samples would normally not be found there (e.g. Culicidae and Cyclopoidea). Furthermore, not all vegetation types were present at all stations during all collecting periods.
Analysis of similarity between sites during each collection revealed the taxa at most stations were different from those of other stations during much of the year (Table 5). Generally, similarity values were highest during April. The highest similarity, 0.727, occurred between Stations 3 and 4 in April. This was followed by Stations 2 and 3 and Stations 1 and 3 with values of 0.621 and 0.600, respectively, also during April. The lowest similarity, 0.100, occurred between Stations 1 and 3 during July. Otherwise, similarity indexes for all station pairs during the duration of the study appeared to fluctuate with no noticeable trends.
Ecological Trends: Boehler Lake appears to fit the general criteria of a dystrophic lake
{Page 27}
as described by Cole (11) and Schalles and Shure (16). These criteria include a shallow basin with stained, acidic waters deficient in calcium and carbonates. The dissolved oxygen concentrations are generally low, probably because of the intense decomposition activity occurring in the detritus on the lake bottom.
The lake also exhibited a high level of turbidity, which may inhibit photosynthetic activity during the warmest months. This
lack of light penetration is most likely due to dissolved matter and to staining of the water. Nutrient concentrations are generally low, which may be indicative of high phytoplankton and vascular plant activity. It is likely that most nutrients are absorbed soon after they become available. The exception to this rapid nutrient absorption was orthophosphate, which remained relatively high during much of the year.
C. punctipennis and chironomid larvae
{Page 28}
seem to dominate microhabitats within the lake. Species similarities between stations within the lake indicate that, during spring, adjacent sampling stations within the lake tend to have somewhat similar community compositions.
Predators dominated in nearly all samples. The collapse of the trophic structure has apparently left this lake with a limited vertebrate population. It appears that trophic structures are built from the bottom up, but maintained from the top down.
There is some evidence to suggest that horizontal movement may be occurring prior to emergence. Autumn species diversity values decreased in the deeper portions of the lake, while those values at shallow water stations showed little change. It is possible that some species moved horizontally from deeper areas to shallower areas of the lake in preparation for emergence.
The rate of detritus accumulation in the lake was not measured. However, based on field observations, it is apparent that the rate of accumulation far exceeds the rate of decomposition. This is similar to the situation described by Horne and Goldman (19) in that nutrient cycling in Boehler Lake appears to be centered around detritus. The deposition of organic matter, combined with migrating and wind-blown soil entering the system, seems to be filling the lake basin.
Boehler Lake is undergoing succession like all natural lentic bodies of water. Because this ecosystem continued to undergo change during the four consecutive seasons it was studied, it is likely that future investigations of this lake will yield different results, supporting its continuing succession. With macroinvertebrate populations at current levels, it is unlikely an aquatic vertebrate population will grow to large numbers because the absence of a significant forage base. In addition, the low DO concentrations will further limit colonization by a fish community. The continued accumulation of soil-building materials throughout the lake basin will keep Boehler Lake shallow and acidic and proceeding toward a terrestrial environment.
Funding to conduct this research came from The Nature Conservancy and The University of Central Oklahoma College of Graduate Studies and Research. J. Hellack and G. Caddell read early drafts of this manuscript and offered helpful suggestions.
| Introduction | Methods | Results and Discussion | References | Top of Page | Table of Contents | Home |
1. Oklahoma Geological Survey. Hydrologic atlas 9 (McAlaster and Texarkana Quadrangles). Norman (OK): University of Oklahoma; 1983.
2. Craven R, Brown B. Benthic invertebrates in Boomer Lake, Payne County, Oklahoma. Southwest Nat 1969; 14:221-230.
3. Craven R, Brown B. Power-plant heated discharge water and benthos in Boomer Lake, Payne County, Oklahoma. J Kans Entomol Soc 1970;43:122-128.
4. Ferraris C, Wilhm J. Distribution of benthic macroinvertebrates in an artificially destratified reservoir. Hydro-biologia 1977;54:169-176.
5. Parrish J, Wilhm J. Relationship between physicochemical conditions and the distribution of benthic macro-invertebrates in Arbuckle Lake. Southwest Nat 1978;23:135-144.
6. Resh V, White D, White S. Lake Taxoma caddisflies (Insecta: Trichoptera): 1. Species present and faunal changes since impoundment. Southwest Nat 1978;23:381-388.
7. Howick G, Wilhm J. Zooplankton and benthic macroinvertebrates in Lake Carl Blackwell. Proc Okla Acad Sci 1984; 64:63-65.
8. Bass D. Colonization and succession of benthic macroinvertebrates in Arcadia Lake, a south-central USA reservoir. Hydrobiologia 1992;242:123-131.
9. Friemuth P, Bass D. Physicochemical conditions and larval Chironomidae (Diptera) of an urban pond. Proc Okla Acad Sci 1994;74:11-16.
{Page 29}
10. Hach Chemical Company. Water and waste-water analysis procedures. Ames (IA): Hach Chemical Co.;1992.
11. Cole G. Textbook of limnology. Prospect Heights (IL): Waveland Press; 1994.
12. Bass D. A simple method to estimate benthic populations of a small woodland stream. Freshwater Invertebr Biol 1985;4:154-157.
13. Shannon C. A mathematical theory of communication. Bell Syst Tech J 1948; 27:379-434.
14. Sorenson T. A method of establishing groups of equal amplitude in a plant society based upon similar content. K Dan Vidensk Selsk Biol Skr 1948;5:1-34.
15. Theinemann A. Die Binnengewasser Mitteleuropas: eine Limnologische Einfuhrung. Binnengewasser 1925;1:1-225.
16. Schalles J, Schure D. Hydrology, community structure and productivity patterns of a dystrophic Carolina bay wetland. Ecol Monogr 1988;59:365-385.
17. DeVito K, Dillion P. Importance of runoff and winter anoxia to the P and N dynamics of a beaver pond. Can J Fish Aquat Sci 1993;50:2222-2234.
18. Merritt RW, Cummins KW. An introduction to aquatic insects of North America. Dubuque (IA): Kendall/Hunt; 1996.
19. Horne A, Goldman C. Limnology. New York (NY): McGraw-Hill; 1994.
Received: January 15, 2001; Accepted: April 17, 2001
{kind=link}
{kind=link}
{kind=link}