
|
|
|
{Page 15}
Department of Biological Sciences, Southeastern Oklahoma State University, Durant,
OK 74701-0609
The Lake Texoma area in Bryan County, Oklahoma, has experienced considerable disturbance as a result of human activity. The area is currently vegetated with a mosaic of forest and grassland. To characterize the vegetation on a 186-ha site in the area adjacent to the lake, we selected and sampled three representative forested areas and two representative grassland areas. Forested areas were sampled by using circular nested quadrats, and grassland areas were sampled by using a 25 cm x 25 cm square quadrat. Two of the three forested areas contained primarily post Quercus stellata (post oak), Ulmus alata (winged elm), and U. americana (American elm), with smaller amounts of Q. marilandica (blackjack oak )and Carya texana (black hickory). These areas were generally similar to other Oklahoma upland forests. The third forested area studied was dominated by winged elm, with few other species present, unlike previously described Oklahoma forest. This is unlike other previously-described Oklahoma forests. Difficulties in identifying major species made quantitative analysis of grassland data impossible. © 2002 Oklahoma Academy of Science
| Introduction | Materials and Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
Lake Texoma, one of the world's largest human-constructed lakes, was completed in 1944. The construction of the lake, and the subsequent recreational use of the adjacent land, has caused disturbance to the vegetation of the area. We studied the vegetation on one parcel of land adjacent to the lake. This 186-ha tract of land, of which we sampled approximately 20%, is located between Johnson Creek and the Roosevelt Memorial Bridge in Bryan County, Oklahoma (lat. 34o 00', long. 96o 30'). We were interested in characterizing the vegetation on this site, and comparing the species composition of the area to that of areas described in previous research on Oklahoma forests and grasslands.
Under Trewartha's system (1), Oklahoma is generally considered to have a subtropical humid climate. The average annual temperature ranges from 15.2oC in the Panhandle to 17.6oC in the extreme southeast. Precipitation varies widely from the drier, western end of the state, which receives about 38 cm per year, to the wetter eastern end, which receives about 125 cm precipitation per year (2). Generally, this precipitation peaks in late spring and again in early fall, with summers being drier. Oklahoma's climate is also influenced by the Bermuda High airmass. This airmass increases the likelihood of dry summers. In years when the high pressure system is displaced to the north and the west, summer droughts are likely (3). A series of drought periods over the course of the last century listed in Rice and Penfound (4) and Tortorelli (5), are 1929-1941 (the most severe), 1949-1954, 1961-1972, and 1975-1982. According to Hoagland (6), these droughts have an average return frequency of 20 years. In addition, there was a localized drought that had effects in south-central Oklahoma from 1984-1986 (5). The region we studied was likely to have been affected by periodic droughts, although after 1948 Lake Texoma may have been a moderating influence.
Bryan County is in south central Oklahoma (4). The mean annual temperature is 16.4oC, with mean June, July, and August temperatures of 25.3oC, 27.9oC, and 27.5oC, respectively. The mean annual precipitation is 108.2 cm, with the months of June, July,
{Page 16}
and August averaging 11.7 cm, 5.8 cm, and 6.6 cm, respectively. May and September show precipitation peaks (7).
Vegetation in Oklahoma varies across the state. A large number of species (as many as 2549 species of vascular plants) are present in the state (6). This is partially because of the variability of habitat type within the state (for example, compare 4 and 8) but also because Oklahoma is in a transition zone between Eastern U.S. and Western U.S. biotas (6).
Historically, Oklahoma has experienced alteration of its native habitats. Timber cutting for lumber or to clear land for agriculture began rather early in the state's history leading Bruner (9) to remark in 1931 that "the best and most accessible timber" had already been removed. However, although many of the original forests have been cut, there has also been forest expansion in Oklahoma. As in much of the grassland region, fire suppression following settlement led to the encroachment of forest on prairie land. In addition, erosion along streambanks following settlement led to expansion of ravines and ravine forests (4).
The Lake Texoma site has experienced considerable disturbance over its history. Although it is currently along the shore of a lake, the vegetation appears to be more similar to upland forest as described by Rice and Penfound (4) than to bottomland or floodplain forest. This is probably because Lake Texoma is an "artificial" lake, created by the Denison Dam (P. Newton, personal communication, 2000). Elevations for the area we studied average 192 m. Originally, the Lake Texoma area was most likely a mosaic of forest and grassland (10). Prior to the 1940s, it was grazed and may have been used for other agricultural purposes. Following development of Lake Texoma, the site has been used mainly for recreation.
The site is located on the boundary of the cross-timbers and the grassland regions (6). Quercus stellata (post oak) and Q. marilandica (blackjack oak) are the major tree species of the cross-timbers region (4) and (11). Johnson and Risser (2) performed ordination analysis on selected forests from Rice and Penfound's upland forest database. They found that Q. stellata and Q. marilandica were the major species of Oklahoma upland forests, but that many sites also had Carya texana (black hickory) and Q. velutina (black oak) present. Their results demonstrated that Q. stellata required more mesic and nutrient-rich conditions than Q. marilandica did, but that the two species are often found together and in association with C. texana and Q. velutina.
The species composition of the forests described in Rice and Penfound (4) are similar to those of the area we studied (leading us to believe that the site contains disturbed upland forest). We conducted this study to determine the species composition of forests present on the site, and to compare our results with published results for less-disturbed Oklahoma forests.
Originally, the study was also to include data from two representative grassland areas at the Lake Texoma site. However, drought conditions in summer 2000 prevented many of the native grass species from flowering, making conclusive identification impossible. We hope to eventually identify these species and conclude the grassland study.
| Introduction | Materials and Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
Study area. The research area is located in Bryan County, between Johnson Creek and Lake Texoma. The site is accessible from US Highway 70 and is bounded on its south side by the highway. The tract of land is 186 ha in size and is managed jointly by the Army Corps of Engineers and the Biology Department of Southeastern Oklahoma State University.
Prior to the development of Lake Texoma, the land in the area was used for agriculture and grazing. Thus it has a disturbance history prior to 1940. After the filling of Lake Texoma, the Corps of Engineers has maintained the land with periodic prescribed burns. Because the land is used for hunting, the Corps of Engineers has also plowed and planted selected areas with Triticum aestivum (wheat), Sorghum bicolor (milo), or Chamaecrista fasciculata (partridge pea) as forage crops to attract game. In addition, they have cut brush and small trees along several paths to permit
{Page 17}
lake access and have cut and sprayed herbicide to control Toxicodendron radicans (poison ivy) along these paths (P. Newton, personal communication, 2000). Pinus echinata (shortleaf pine), P. taeda (loblolly pine), and Elaeagnus angustifolia (Russian olive) have been planted as part of botany classwork or research projects in the past. Also, in the summer of 2000, Alnus maritima (seaside alder) was planted along the lakeshore as part of a research project but appears not to have survived. We selected our sampling areas to avoid these areas of recent disturbance.
The Lake Texoma site is prone to flooding and to fluctuations in the elevation of the lake. In 1957 and again in May 1990, there were large floods that covered the entire area (P. Newton, personal communication, 2000). In spring 2001, several low-lying areas of the site were submerged for about a month.
In the summer of 2000, we began our survey of the area. We selected three forested areas that were representative of forests on the site as a whole and two grassland communities representative of grassland communities on the site.
Sampling Techniques. In the forested areas, we used the circular nested quadrat technique (12). Three ropes of varying lengths were attached to a stake. The stake represented the center of the sampling plot, and the quadrat was the circle scribed by each rope as it was rotated about the central point. The three quadrats had different areas: .04 ha for trees, .01 ha for saplings, and .001 ha for seedlings. For the purposes of this study, a tree was defined as any tree with a diameter at breast height (DBH) in excess of 9 cm; a sapling was a tree with a DBH less than 9 cm but taller than 20 cm; and a seedling was a tree less than 20 cm tall. Only trees were included in the sampling; there is woody brush such as Symphoricarpos orbiculatus (buckbrush) and Smilax spp. (greenbriar) present in the understory. The number of quadrats sampled per forest were proportional to the size of the area; 12 quadrats each were sampled in the lake and ravine areas whereas 9 quadrats were sampled in the smaller elm grove area. Voucher specimens of flowering individuals were collected and were deposited in Southeastern Oklahoma State University's (SOSU) herbarium. Nomenclature follows Tyrl and Waterfall (13); Little (14) was used as an aid for field identification.
We analyzed the circular nested quadrat data following the procedure outlined in Curtis and Cottam (12). Relative dominance (based on basal area), relative density, and relative frequency were calculated for each tree species at each sample site. Values were calculated as percentages and used to compute an importance value (as an average of the three values) for each tree species in each forested area (See Curtis and Cottam (12), for more details). The importance value (IV) is comparable to the "importance percentage" of Rice and Penfound (4).
We computed relative density and relative frequency values for sapling and seedling layers but not relative dominance. We averaged the relative density and relative frequency to compute an importance value for seedlings and salings.
Additionally, we compared species composition of three woodland areas. We computed Sorenson's index of similarity (15) lake and ravine areas, for lake and elm grove areas, and for ravine and elm grove areas for tree, the sapling, and the seedling strata. We also computed Shannon diversity indices and evenness (15) for each stratum of each area.
To sample the grassland areas, we used the quadrat frequency method (16). We worked along 5 north to south transects in the smaller area and 13 north to south transects in a larger area. Each transect was separated by approximately 10 m. We sampled up to 12 quadrats per transect, depending on transect length. Quadrat samples were spaced at approximately 10 m intervals. At each sampling point, stratified random sampling was used to locate the exact point of sampling. A 25 cm x 25 cm sampling frame was used as the quadrat. For each sample, the species present in the quadrat were recorded, allowing development of a species list for the grassland area and calculation of a
{Page 18}
relative frequency of each species. Voucher specimens of flowering species were collected, identified, and deposited in SOSU's herbarium. Because of a severe drought in summer 2000, not all species on the site successfully flowered. As a result it was not possible to identify some of the major grass species on the site. This made quantitative analysis of the data impossible. We hope to continue this project and identify the major warm-season grass species.
| Introduction | Materials and Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
Two of the three forested areas showed compositional similarities. Both were dominated by Q. stellata. The third site, however, was different in appearance (it had almost no understory), in diversity, and in species composition [it was almost entirely composed of many, closely spaced, fairly small U. alata (winged elm)], and in level of diversity. We referred to the three areas as "ravine area," "lake area" and as "elm grove." (However, the "ravine area" is not a true ravine; we called it that because a dry streambed ran through the area. The total elevation difference in the area was less than 5 m). The lake and ravine areas were more similar to each other in terms of tree, sapling, and seedling composition than either was to the elm grove.
The three sites differed in species richness and diversity. The ravine area was the most rich, having the greatest numberof species in the tree stratum (12), whereas the lake area had 8 species present, and the elm grove only 5. Diversity indices for each site are shown in Table 1. The lake area had a Shannon diversity of 0.783 with an evenness of 82%. The ravine area was slightly more diverse, with Shannon diversity of 0.838, but less even, with evenness of 75%. However, the elm grove area had the lowest diversity, with Shannon diversity of only 0.073 and an evenness of 10%, reflecting the high abundance of Ulmus alata.
In the ravine area, the tree stratum was dominated by Q. stellata, with a relative importance value of 30%; U. alata, with a relative importance value of 16%; Q. marilandica, with a relative importance value of 10%, and U. americana (American elm) with a relative importance value of 10%. Other species present, in descending order of importance, were C. texana, Q. velutina, Morus alba (white mulberry), Fraxinus americana (white ash), Maclura pomifera (Osage orange), and minor quantities of Sideroxylon lanuguinosum (chittamwood), Carya illinoense (pecan), and Celtis laevigata (sugarberry) (see Table 2). The herbaceous layer contained a variety of small forest plants as well as T. radicans, Smilax spp., and Rubus sp. (blackberries). This area was most similar to the Quercus stellata—Ulmus alata forest association described by Hoagland (6) or to Cross-Timbers forest (11). The understory, however, did not contain the same
{Page 19}
species as described by Hoagland (6), nor did it contain species typical of Cross-Timbers forest understory (11). Both of these types of areas contain understory that has a component of prairie grasses, including Andropogon gerardii (big bluestem) and Schizachyrium scoparium (little bluestem). It seems likely that past disturbance in the area could have led to changes in the understory – most of the species currently present are species that come in after disturbance.
The lake area had a similar species composition and abundance. This site was dominated by Q. stellata [importance value (IV) = 28%], winged elm (IV=14%), and U. americana (IV=11%). There were smaller amounts of Q. marilandica, C. texana, Maclura pomifera, S. lanuginosum and Morus alba present. This site lacked Q. velutina and F. americana. Again, this area was most like the Quercus stellata—Ulmus alata forest type described by Hoagland (6). Again, the understory vegetation showed little relationship with the understory described in either Hoagland (6) or (11); this is probably also a result of past disturbance.
Both of these forested areas were similar in their tree-stratum species com-
{Page 20}
position, with a Sorenson coefficient of 76.1% (see Table 3 for Sorensen coefficients). The two areas were also similar in general appearance, with many of the same species present in the ground layer, including Carex spp. (sedges), T. radicans, Rubus sp., and Chasmanthium latifolium (inland sea-oats). The elm grove area, however, was extremely different in its species composition and appearance. This forested area was almost entirely dominated by U. alata (IV=86%), with much smaller quantities of Diospyros virginiana (persimmon), Sophora affinis (sophora), U. americana, and C. laevigata. The elm grove area had a low similarity with either the lake or the ravine area. The tree stratum for the elm grove has a Sorenson similarity of 18.2% with the lake area and 16.7% with the ravine area.
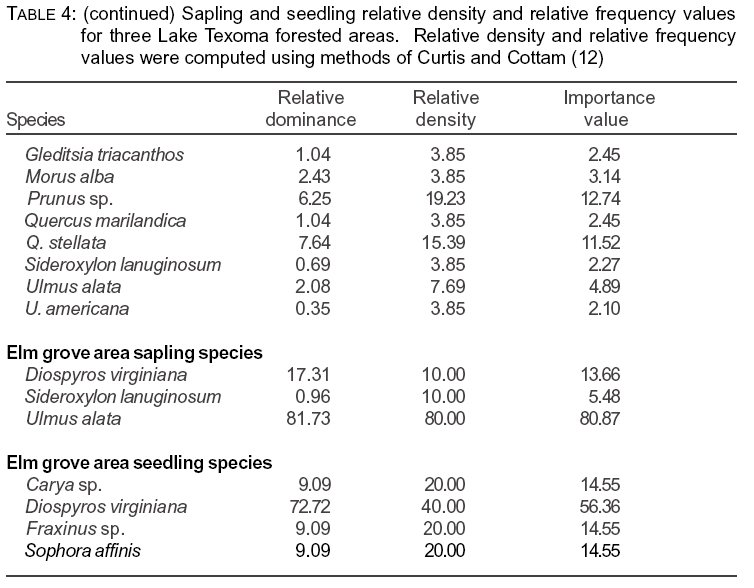
One concern when studying forested areas is whether the area is undergoing change or whether the tree species present are successfully reproducing themselves. We compared tree, sapling, and seedling strata of these three forested areas. The sapling strata contain both younger examples of trees (such as Q. stellata) as well as trees that tend to be smaller than 9 cm DBH at maturity (for example, S. lanuginosum). Importance values for saplings and seedlings are presented in Table 4.
In the ravine area we found the Fraxinus sp. (it was difficult to determine if most were F. americana or F. pennsylvanica); saplings had the highest relative importance value (44.86%), followed by S. lanuginosum (13.65%), and U. alata (10.53%). Carya texana (IV=8.37%) and Q. stellata (IV=6.22%) were present in lower abundance. The seedling stratum showed a different pattern, with C. texana (IV=28.89%) being the leading species present, with U. alata (IV=16.65%) and Fraxinus sp. (IV=11.11%) also having high abundance.
In the lake area the most abundant species in the sapling layer were Q. stellata (IV=35.14%), followed by C. texana (IV= 11.57%), U. alata (IV=11.15%), and Fraxinus sp. (IV=9.81%). The seedling stratum had high abundance of C. texana (IV=40.69%), with lower abundance of Prunus sp. (cherry) (IV=12.74%), Q. stellata (IV=11.52%) and Cornus drummondii (rough-leaf dogwood) (IV=10.44%).
In the elm grove area U. alata was the major sapling/small tree, with an importance value of 80.9%. The only other species present in the sapling stratum were D. virginiana (IV=13.6%) and S. lanuginosum (IV=5.46%). The seedling stratum, however, contained no individuals of U. alata, and was dominated by D. virginiana (IV=56.4%) with smaller numbers of Sophora affinis, Fraxinus sp, and C. texana.
| Introduction | Materials and Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
The three forested areas were selected to be representative of the forest vegetation on the site as a whole. Two of these areas (lake and ravine) were generally similar. Furthermore, they were similar in species composition to the upland oak-hickory forest type described by Rice and Penfound (4) and to the Quercus stellata—Ulmus alata forest type described by Hoagland (6). The understory of these sites did not reflect the understory described in Hoagland (6) or Hoagland (11) and may be showing the result of past disturbance or grazing. Still, the sites do resemble oak-hickory upland forest.
However, the third site, the elm grove, was markedly different from the other two Lake Texoma forests we studied and did not
{Page 21}
{Page 22}
correspond well to any of the forest types previously described for Oklahoma (4, 6, 18). It is probably a forest that developed following some disturbance at the site; Baker (17) has described patterns of invasion of disturbed forested areas and U. alata is one of the species capable of becoming invasive.
The lake and the ravine areas were dominated by Q. stellata. The two areas were generally similar in species composition. In the ravine area, Q. stellata was the most important species, followed by U. alata, Q. marilandica, and U. americana. Other species typical of Oklahoma upland forests were present in lower abundance, specifically Q. velutina and C. texana. In the lake area, Q. stellata had the highest importance value, followed by U. alata, U. americana, and Q. marilandica. Carya texana was a minor species (as it was in the ravine area), but black oak was not present on this site.
The lake and the ravine areas are similar in species composition of the tree stratum. However, the ravine area has a larger number of species present (12) than the lake area (8). Specifically, the lake area lacks Q. velutina, C. illinoense, Celtis laevigata, and Fraxinus americana. Quercus velutina has higher moisture requirements than either Q. stellata or Q. marilandica (2). Carya illinoense and Celtis laevigata are listed among typical species of bottomlands (8), suggesting they have higher moisture requirements. It seems that the ravine area is most likely moister than the lake area.
Although these two sites displayed some differences, they were both generally similar to the oak-hickory types upland forest described by Rice and Penfound (4) or to the Quercus stellata—Ulmus alata forest type described by Hoagland (6). The major species of upland oak-hickory forest, according to Rice and Penfound (4), were Q. stellata, Q. marilandica, Q. velutina, C. texana, and U. alata. All of these species were present at the ravine area and all except black oak were present at the lake area. However, there are a number of species listed by Rice and Penfound (4) as typical of upland oak-hickory forest that are not present at the Lake Texoma sites. These species are Carya tomentosa (mockernut
{Page 23}
hickory), Quercus shumardii (Shumard oak), Quercus alba (white oak), Quercus muhlenbergii (chinquapin oak), and Acer saccharum (sugar maple). Many of these species tend to have higher soil moisture requirements than Q. stellata and Q. marilandica, and it is possible that these species are absent because the Lake Texoma sites are too dry for them. The Lake Texoma sites also differed from Rice and Penfound's (4) description of upland oak-hickory forest because they had some species that Rice and Penfound (4) do not list. The most important of these species is U. americana. However, the ravine area also had F. americana, Morus alba, C. laevigata, Maclura pomifera, and C. illinoensis present in low abundance. Ulmus americana is known to be a species that can grow aggressively on disturbed areas (18). Morus alba is an exotic species but is spread by birds, as is the native species C. laevigata. It appears that the two main Lake Texoma forest sites we studied were somewhat similar to dry upland oak-hickory forests as described in Rice and Penfound (4) and Hoagland (4) but show effects of disturbance.
Although the tree strata of the ravine and lake areas were generally similar (and were representative of most of the forest on the site), the third area we studied was markedly different. We referred to the site as the elm grove area because of the near-total dominance of Ulmus alata. This site had an importance value of 85.7% for U. alata, with very few other species present. Two of the forests described in Rice and Penfound (4) were dominated by U. alata, but they also had other typical species of upland forest (such as Q. stellata and Q. marilandica) present. Thus, the elm grove forest at the Lake Texoma site is different from previously described Oklahoma forests. We think that this unusual species composition may result from a combination of soil characteristics (The Bryan County Soil Survey (10) describes the site as having a clay pan present) and possibly of disturbance. Baker (17) describes "pure stands" such as the one we observed as being a development after disturbance of an area. However, when we cored three of the larger trees on the site, we found them to be 18 years old – an establishment time that does not correlate well with known past disturbances. Current research is ongoing to determine why this area (and two other forested areas on the site) have such high dominance of U. alata. In spring 2002, we did notice heavy herbivore damage to the winged elms in the elm grove area.
We were also interested in determining whether the site was likely to undergo a change in species composition over time. We determined this by examining data on seedlings and saplings. It appears that the oaks are not completely successful in their reproduction in the ravine and lake areas. In the ravine area the most abundant saplings were Fraxinus sp., S. lanuginosum (actually a small tree that is unlikely to attain canopy height) and U. alata. There were a few saplings of Q. stellata. In the seedling stratum, Carya texana is dominant, followed by Fraxinus sp. There is a higher density of C. texana saplings and seedlings than there are trees in the ravine area, suggesting that recent reproduction of C. texana has been unusually high or that recruitment of seedlings is poor. The high density of Fraxinus seedlings suggests that the site may be shifting over time to be more of a closed-canopy forest. However, the Fraxinus seedlings and saplings present may not survive to tree status. Also, in Spring 2001 and 2002, the ravine area was burned, which likely had an effect on the seedling layer.
Quercus stellata was more successful in reproduction at the lake area. It was the species with highest importance value in the sapling stratum, followed by C. texana and U. alata. In the seedling layer, C. texana was the species with highest density, followed by Prunus sp., Q. stellata, and C. drummondii. It appears that the lake area is more likely to remain an oak-hickory type forest than the ravine area is. The high reproduction of Q. stellata is interesting in light of the presence of deer on the site, and the fact that deer feeding stations have been set up near the lake area forest. Many forests with high deer density are experiencing effects on their seedling and sapling layers. Apparently this is not the case at the Lake Texoma site. Possibly, hunting pressure is sufficient to keep the size of the deer herd under control.
{Page 24}
The elm grove area seedling layer shows the greatest variability. In the sapling layer U. alata was still the dominant species, but there were no U. alata seedlings found in any of the samples taken. Germination of U. alata seeds is generally high (Bannister, unpublished data, found germination percentages as high as 82% and Kirby (19) as high as 96%). However, the seeds have poor survival in the seed bank (Bannister, unpublished data) and Kirby (19) shows a fairly high mortality of U. alata seedlings. There were, in fact, very few seedlings of any species (persimmon was the species having the greatest number of seedlings). The elm grove area also has very little ground cover. The absence of seedlings and ground cover may be related to the clay pan soils, which tend to be very wet in early spring and intensely droughty in late summer.
The authors thank the staff of the Texoma-area Corps of Engineers for permission to visit and use the Lake Texoma site for research. We especially thank Philip Newton for his discussion of the history of Lake Texoma and the research area. Facilities for identification of plant species and soils lab equipment were provided by the Biological Sciences Department of Southeastern Oklahoma State University.
| Introduction | Materials and Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
1. Trewartha GT. An introduction to climate. 4th ed. New York: McGraw-Hill Book Co., Inc.: 1968. 408 p.
2. Johnson FL, Risser PG. Some vegetation environment relationships in the upland forests of Oklahoma. J Ecol 1972; 60:655-663.
3. Cooter E. General climatology in Oklahoma. In: Tortorelli RL, editor. Floods and droughts: Oklahoma, national water summary 1988-89. US Geological Survey, Water Supply Paper 2375; 1991; 451-452.
4. Rice EL, Penfound WT. The upland forests of Oklahoma. Ecol 1959; 40:593-608.
5. Tortorelli RL. Floods and droughts: Oklahoma, national water summary 1988-89. US Geological Survey, Water Supply Paper 2375; 1991. 512 p.
6. Hoagland B. Vegetation of Oklahoma: landscape mapping and conservation planning. Southwestern Naturalist 2000; 45: 385-420.
7. Oklahoma weather [online]. Available: http://www.weather.ocs.ou.edu . (Accessed on August 28, 2001; as of 22 July 2002, website is no longer accessible)
8. Brabander JJ, Masters RE, Short RM. Bottomland hardwoods of eastern Oklahoma: a special study of their status, trends, and values. Oklahoma City (OK): Oklahoma Department of Wildlife Conservation 1985; 144 p.
9. Bruner WE. The vegetation of Oklahoma. Ecol Mono 1931;1:99-188.
10. United States Department of Agriculture. Soil survey of Bryan County, Oklahoma. USDA Soil Conservation Service and Oklahoma Agricultural Experiment Station. 1978. 110 p. Available from Bryan County OSU extension agency.
11. Hoagland BW, Butler IH, Johnson FL, Glenn S. The cross timbers. In: Anderson RC, Fralish JS, Baskin JM, editors. Savannas, barrens, and rock outcrop plant communities of North America. New York: Cambridge University Press; 1999; 231-245.
12. Curtis JT, Cottam G. Plant ecology workbook. Minneapolis (MN): Burgess Publishing Company; 1962. 193 p.
13. Tyrl RJ, Waterfall UT. Identification of Oklahoma plants : a taxonomic treatment comprising keys and descriptions for the vascular plants of Oklahoma and keys to the flora of Oklahoma, 6th edition. Stillwater (OK): Flora Oklahoma, Inc.; 2000. 246 p.
14. Little EL Jr. Forest trees of Oklahoma. Oklahoma City (OK): Oklahoma Forestry Services Publication no. 1; 1989. 205 p.
15. Magurran A. Ecological diversity and its measurement. Pinceton (NJ): Princeton University Press;1988. 179 p.
16. Sorells L, Glenn S. Review of sampling techniques used in studies of grassland plant communities. Proc Okla Acad Sci 1991;71: 43-45.
17. Baker FS. Principles of silviculture. New York: McGraw-Hill Book Co.; 1950. 414 p.
18. Barnes BV, Wagner WH Jr. Michigan trees. Ann Arbor (MI): University of Michigan Press; 1996. 383 p.
19. Kirby BW Responses of winged elm (Ulmus alata Michx.) to various methods of herbicide treatment. [MS Thesis]. Oklahoma State University; 1962. 54 p. Available from: OSU Library.
Received: December 20, 2001; Accepted: June 24, 2002.