
|
|
|
{Page 73}
Department of Zoology and Oklahoma Cooperative Fish and Wildlife Research Unit, United States Biological Survey, Biological Resources Division, Oklahoma State University, Stillwater, OK 74078
1Current address: Mississippi Museum of Natural Science, 2148 Riverside Drive, Jackson, MS 39202-1353
Raccoons and opossums use similar habitats in North America and share food preferences. We conducted a 1-year raccoon-removal study at the Cross Timbers Experimental Range (CTER), Stillwater, Oklahoma, and compared diets of opossums in nonremoval and removal areas to assess the potential competitive influence of raccoons on opossum diets. A total of 75 scats were collected from 65 unique opossums captured in traps from nonremoval (n = 28) and raccoon-removal (n = 47) areas. Major food items were invertebrates, soft mast, grass, and woody plant leaves. Results from c2 analysis suggested a lack of differences in major food items between opossum scats collected in non-removal and raccoon-removal pastures for the duration of the study. The lack of differences in diets of opossums between areas suggested minimal interspecific food competition between raccoons and opossums at the spatial and temporal scale of this study. Interspecific competition for food may not occur at CTER because of differential habitat selection or diet partitioning between the two species. © 2002 Oklahoma Academy of Science
| Introduction | Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
Food habit studies describe an animal's use of habitat and surrounding biota based on prey selection (1). Studies examining interspecific competition in relation to food habits among rodents and marsupial carnivores have found larger species competitively exclude smaller competitors through exploitative or interference competition (2,3).
The food habits of mesocarnivores, such as raccoons (Procyon lotor) and opossums (Didelphis virginiana), have been investigated for comparative purposes without determining interspecific interactions (1,4,5). Opossums and raccoons consume similar prey items (1), thereby increasing the potential for interspecific interactions that may lead to exploitative and/or interference competition (6). They occur in similar habitat throughout their range, and habitat-use overlap has been recorded as high as 95% (7). Ladine (8) found that raccoons and opossums used available habitat during different hours. On the basis of the absence of dual species captures at double-trap sites, the presence of larger raccoons may force smaller opossums to forage at different times, suggesting competitive exclusion pressures for habitat use potentially affecting opossum prey selection (6).
Several studies have determined that opossums and raccoons consume similar prey items, but have slight differences in preference (1,4,5,9,10,11). These studies analyzed scat and/or digestive tract (stomach, small intestine, and colon) contents for diet determination. Opossums are omnivorous, but prefer small-animal prey, such as insects (1,9,10; Table 1). Carrion, rodents, and other opossums are the most common mammalian prey (5,11). Plants
{Page 74}
constitute a large portion of opossum diets (1,5,9,10). Raccoons are also omnivorous generalists, consuming insects, but eat hard and soft plant matter more frequently (1,4). They prefer fruits such as grapes (Vitis spp.), apples (Malus spp.), and persimmons (Diospyros spp.), as well as acorns (Quercus spp.) and corn (Zea mays). Aquatic invertebrates are consumed in greater numbers than are vertebrates. Rodents are the most abundant preferred mammals eaten. Raccoons also eat bird and reptile eggs (e.g., Anas platyrhynchus, Chelydra serpentina, Alligator mississippiensis) (1,12,13).
Manipulation of the natural system by adding or removing individuals is necessary to understand whether resources are partitioned and whether competition is involved in structuring a community (14). To determine whether competition exists between raccoons and opossums, we conducted a raccoon-removal study to assess the effects of raccoon absence on opossum ecology at the Cross Timbers Experimental Range (CTER) in Payne County, Oklahoma. Specifically, the objectives of this study were to identify changes in opossum diet following raccoon removal. We predicted opossums living in raccoon-removal sites would exhibit a diet shift because of the release associated with removal of a larger, dominant competitor.
| Introduction | Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
CTER is located 11 km southwest of Stillwater, Payne County, Oklahoma (36°02'40" to 36°04'20"N, 97°09'30" to 97°11'39"W). It is a 712-ha area characterized by an overstory dominated by post oak (Quercus stellata), blackjack oak (Q. marilandica), and American elm (Ulmus americana) interspersed with invading eastern redcedar (Juniperus virginiana). Little bluestem (Schizachyrium scoparium), indian-grass (Sorgahastum nutans), switchgrass (Panicum virgatum), grama grasses (Bouteloua spp.), purpletop (Tridens flavus), ragweed (Ambrosia spp.), and buckbrush (Symphoricarpos orbiculatus) are prevalent in the understory (15). This area has been studied since 1983 to determine the effects of different methods of rangeland manage-ment on vegetation (15-17), livestock production, and wildlife. Four distinct habitat types exist at CTER as a result of the treatments (e.g., burning, herbicide) that differentially influenced conifer and deciduous trees and shrubs (D. M. Engle, Department of Plant and Soil Sciences, Oklahoma State University, personal communication). These habitat areas were classified by Levesque (18) as redcedar forest, derived grassland, mixed-brush community, and mature oak forest.
{Page 75}
Our study site was composed of 130-ha removal and non-removal areas. Each area contained four trapping grids and was a habitat mosaic produced by the same four land management prescriptions. We trapped bi-monthly in both areas from January 2000 to January 2001 for 10-day periods. Animals were trapped using Tomahawk live traps (Tomahawk Trapping Company, Tomahawk, WI) baited with sardines. Each trapping grid contained eight traps, with two sets of three traps 300 m apart along parallel transects spaced at 200 m intervals, and two interior traps placed 180 m diagonally from the corners (18). Eight traps were placed between nonremoval and treatment grids, around the perimeter of the treatment grids, as buffers to minimize raccoon immigration into treatment grids.
We checked traps daily and rebaited every 5 days and as needed during the 10-day trapping sessions. We chemically immobilized (8.0-mg/kg Telazol®, A. H. Robins Company, Richmond, Virginia, USA), ear tagged, sexed, and took morphological measurements on opossums and raccoons. We determined opossum age using body mass (> 0.9 kg = adults, < 0.9 kg = juveniles). Female opossums in all pastures weighing > 0.9 kg were fitted with a radiocollar for tracking purposes. Once handling procedures were completed, we released opossums at their point of capture. We transferred raccoons from removal areas by truck and released them at a location, approximately 11 km east of the removal grid.
Opossum diet was evaluated from the collection and analysis of scat found at trap sites from both removal and nonremoval grids. We collected scats only from opossums that had deposited scats in traps, and we assumed opossums captured in a particular area fed in that area. Date of collection, animal identification number, sex, and location of collection (area type and trap site) were recorded. Scats collected from recaptured individuals on different days were considered separate samples. Collected scats were stored in a freezer until the end of each trapping period when they were processed for analysis.
Frozen scats were sieved (USA Standard Testing Sieve, No. 30, Opening 600 mm Tyler equivalent, W.S. Tyler Incorporated, Mentor, OH 44060) under warm water to remove soil particles. Wet, digested food items were air-dried in an oven at 60°C for several hours. Dried particles were separated with tweezers in a plastic petri dish and were then placed under a dissecting scope (50X magnification) for identification. All prey items were identified, regardless of abundance per scat, and were categorized as mammal, bird, amphibian, reptile, invertebrate, plant mast (mast = fruit, soft and hard), leaf (grasses, woody plants [hardwoods and eastern redcedar], forbs), groom hair, root, or moss. Data were recorded for each scat as presence/absence of individual food categories.
Food items were summarized based on frequency of occurrence (19) for scats collected in raccoon-removal and non-removal grids. Chi-square analysis was used to compare distributions of food items in raccoon-removal and non-removal grids during the entire study. Frequency of occurrence data were converted to proportional frequency to calculate Levins' index (20):
B = (Σpi2)-1
where pi is the proportional use of a food item relative to other food items. B ranges from 1 to n (n = total number of food item categories; mammal, bird, etc.) and was used to calculate diversity of diets annually and seasonally for raccoon-removal and non-removal areas. Diversity was standardized to a scale of 0.0 to 1.0 by using Hurlbert's method (21):
Bs = (B-1)/(n-1)
where B = Levins' measure of diversity and n = number of food categories. Diversity indexes and seasonal comparisons of percent occurrence were not statistically analyzed and are provided for descriptive purposes only.
{Page 76}
| Introduction | Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
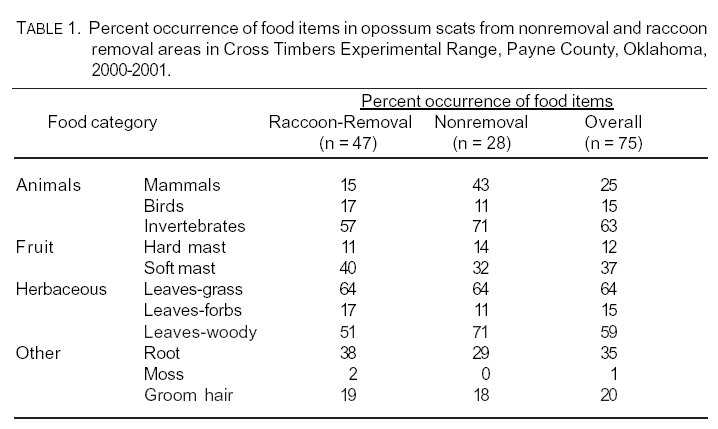
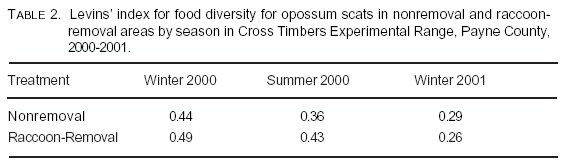
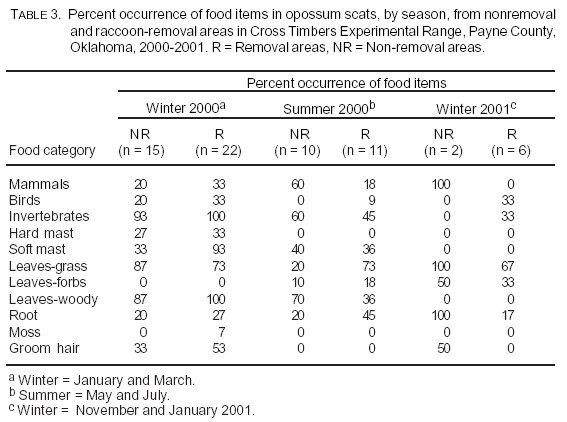
Frequency of occurrence of food categories did not vary between raccoon-removal and nonremoval areas (c212 = 0.99, P > 0.05; Table 1). Levins' index for food diversity for scats across all seasons was Brs = 0.56 and Bnrs = 0.45 in raccoon-removal and nonremoval areas, respectively. Food diversity was slightly higher for scats found in raccoon-removal grids during winter and summer 2000 (Table 2). The increase in soft mast was mainly due to a high occurrence of fruit from eastern redcedar. Percent occurrences of food items varied because of season (Table 3).
| Introduction | Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
We had predicted that food items in opossum scats from raccoon-removal grids would vary from those in nonremoval grids because of competitive release. Levins' indexes for raccoon-removal pastures were higher for all seasons except winter 2001, which had small samples. Raccoon-removal scats were slightly more diverse in food type rather than nonremoval scats. However, these differences may be caused by differences in food availability in nonremoval and treatment groups, individual opossum preferences, and uneven sample sizes rather than competitive release.
{Page 77}
Competition for food resources between raccoons and opossums may be minimized if they partition habitats. Opossum capture rates were higher in oak forest areas at CTER, whereas raccoons were more likely to be captured in cedar forests (18,22). Opossum scats from raccoon-removal areas had a higher percent occurrence of cedar fruit. This finding coincides with analyses regarding microhabitat selection of opossums at CTER, where opossums shifted toward cedar habitat during the study period in raccoon-removal areas (22) and hints at a subtle diet shift in the absence of raccoons.
Opossums at CTER largely consumed invertebrates (insects), soft mast (e.g., eastern redcedar), and leaves from grasses and woody plants (Quercus spp). Variation in percent occurrence of food items at CTER was likely due to differences in habitat, food availability, timing of collection, or sample size. The occurrence of mammal remains in opossum scats at CTER was similar to the occurrence of mammals in scats from central Missouri (10), Kansas (11), and eastern Texas (1). Parts of CTER resemble the Post Oak Woods area of eastern Texas, where post oak and black jack oak are the dominant tree species. Occurrence of mammals in CTER scats was most different from those from southern Illinois (5).
Invertebrate prey, such as insects, is very important in opossum diets. Occurrence of insects in CTER scats were most similar to scats collected in central Missouri (10) and Kansas (11), slightly lower than scats in southern Illinois (5), but least similar to opossum scats in eastern Texas (1). Raccoon-removal scats closely resembled southern Illinois scats in occurrence of birds (5), certainly a matter of chance availability. CTER scats did not contain amphibians. Diet analysis studies in eastern Texas (1) and southern Illinois (5) were the only studies to have an amphibian occurrence and at a relatively low occurrence to other food items. Finally, plants were common in scats from all other studies. Opossum scats from CTER most resembled scats in central Missouri (10) and east Texas (1).
The lack of evidence for interspecific competition between raccoons and opossums at CTER would be better supported if food abundance and availability had been concurrently assessed on areas in non-removal and raccoon-removal grids. Availability data would have allowed assessment of opossum diet selection, not merely use, in the two grids. In addition, the diversity of raccoon and opossum diets makes it more difficult to detect diet partitioning than in studies of obligate carnivores (2).
This research was conducted under Animal Care and Use Protocol 723 at Oklahoma State University. Logistical support was provided by D. M. Leslie, Jr., and the Oklahoma Cooperative Fish and Wildlife Unit (U.S. Geological Survey-Biological Resources Division, Oklahoma State University, Oklahoma Department of Wildlife Conservation, and Wildlife Management Institute, cooperating). Funding was provided by the Department of Zoology and Department of Plant and Soil Sciences at Oklahoma State University. We thank the many graduate and undergraduate students who helped with field work.
| Introduction | Methods | Results | Discussion | References | Top of Page | Table of Contents | Home |
1. Wood JE. Food habits of furbearers in the upland post oak region of Texas. J Mamm 1954;35:406-414.
2. Jones ME, Barmuta LA. Niche differentiation among sympatric Australian dasyurid carnivores. J Mamm 2000; 81:434-447.
3. Dickman CR. An experimental study of competition between two species of dasyurid marsupials. Ecol Monographs 1986;56, 221-241.
4. Hamilton WJ Jr. The food of the opossum in New York State. J Wildl Man 1951;15:258-264.
5. Stiegltiz WO, Klimstra WD. Dietary pattern of the Virginia opossum, Didelphis virginianus Kerr, late summer-winter, southern Illinois. Trans Ill Acad Sci 1962;55:198-208.
6. Ladine TA. Activity patterns of cooccurring populations of Virginia opossums (Didelphis virginiana) and rac-
{Page 78}
coons (Procyon lotor). Mammalia 1997;61:345-354.
7. Kissell RE Jr., Kennedy L. Ecological relationships of cooccurring popu-lations of opossums (Didelphis virginiana) and raccoons (Procyon lotor) in Tennessee. J Mamm 1992;73:808-813.
8. Ladine TA. Ecology of cooccurring populations of Virginia opossums (Didelphis virginiana) and raccoons (Procyon lotor) [Ph.D. dissertation]. Memphis (TN): Univ Memphis; 1995. 90 p. Available from: UofM Library.
9. Lay DW. Ecology of the opossum in eastern Texas. J Mamm 1942;23:147-159.
10. Reynolds HC. Some aspects of the life history and ecology of the opossum in central Missouri. J Mamm 1945; 26:361-379.
11. Sandidge LL. Food and dens of the opossum (Didelphis virginiana) in northeastern Kansas. Trans Kans Acad Sci 1953;56:97-106.
12. Kaufmann JH. Raccoon and allies. In: Chapman JA, Feldhamer GA, editors. Wild mammals of North America: biology, management, and economics. Maryland: Johns Hopkins University Press; 1982. p 567- 585.
13. Sanderson GC. Raccoon. In: Novak M, Baker JA, Obbard ME, Malloch B, editors. Wild furbearer management and conservation in North America. Ontario (CAN): Ontario Trapper's Assoc; 1987. p 487-499.
14. Schoener TW. Resource partitioning in ecological communities. Science 1974; 185:27-39.
15. Ewing JH, Stritzke JF, Kulbeth J. Vegetation of the Cross Timbers Experimental Range, Payne County, Oklahoma. Stillwater (OK): Research Report P-586, Oklahoma Agricultural Experimental Station 1984.
16. Engle DM, Stritzke JF, McCollum FT. Vegetation management in the Cross Timbers: response of understory vegetation to herbicides and burning. Weed Tech 1991;5:406-410.
17. Stritzke JF, Engle DM, McCollum FT. Vegetative management in the Cross Timbers: Response of woody species to herbicides and burning. Weed Tech 1991;5:400-405.
18. Levesque LP. Effect of land-use manipulations on habitat associations and demography of mesocarnivores in the Cross Timbers ecoregion of Oklahoma [MS Thesis]. Stillwater (OK): Oklahoma State Univ; 2001. 102 p. Available from OSU Library.
19. Litvaitis JA, Titus K, Anderson EM. Research and management techniques for wildlife and habitats. Measuring vertebrate use of terrestrial habitats and foods. In: Bookhout TA, editor. 5th ed. Bethesda (MD): Wildlife Society; 1996. p 254-274.
20. Levins R. Evolution in changing environments. Princeton (NJ): Princeton University Press; 1968.
21. Krebs CJ. Niche overlap and diet analysis. In: Krebs C, editor. Ecological methodology. New York, (NY): Harper and Row; 1989. 654 p.
22. Ginger SM. Effect of species removal on carnivore community habitat use in a managed cross timbers ecosystem [MS Thesis]. Stillwater (OK): Oklahoma State University; 2001. 127 p. Available from: OSU Library.
Received: May 15, 2002; Accepted: August 29, 2002